Чем представлена транспортная система клетки
• Клетки эукариот обладают развитой системой внутренних структур, окруженных мембранами, которые называются органеллами
• Каждая органелла обладает уникальным составом (глико)протеинов и (глико)липидов и выполняет определенный набор функций
• Каждая органелла содержит один или несколько компартментов, ограниченных мембранами
• Органеллы выполняют свои функции автономно или в группах
• При эндоцитозе и экзоцитозе переносимые белки (белки карго) транспортируются между компартментами через транспортные везикулы, которые образуются отпочковыванием от поверхности органеллы и затем сливаются с мембраной-мишенью акцепторного компартмента
• Транспортные везикулы могут избирательно включать переносимый материал и исключать те компоненты, которые должны оставаться в органелле, из которой везикулы образовались
• Селективное включение в везикулы обеспечивается сигналами, присутствующими в первичной структуре белка или в углеводной структуре
• Транспортные везикулы содержат белки, которые направляют их к местам назначения и связывания. В дальнейшем везикулы сливаются с акцепторным участком мембраны
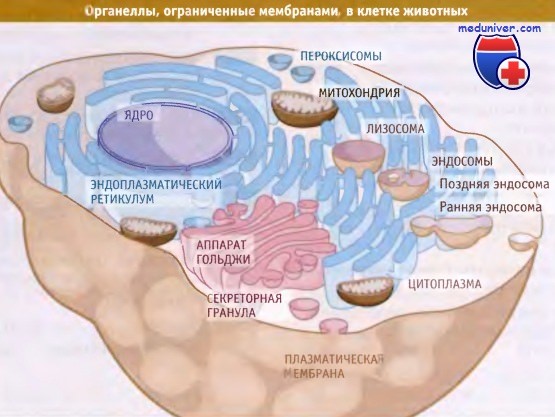 Компартменты, ограниченные мембранами в типичной клетке животных.
Компартменты, ограниченные мембранами в типичной клетке животных.
Одной из характерных особенностей эукариотической клетки является присутствие в ней развитой системы внутренних структур, окруженных мембранами, которые называются органеллы. Для клеток эукариот характерно присутствие мембран, разделяющих их внутреннее содержимое на функционально-различные компартменты, тогда как все клетки живых организмов обладают наружной двухслойной мембраной.
Одно из преимуществ компартментализации состоит в том, что клетка имеет возможность создавать необходимое окружение для выполнения функций, требующих определенного химического состава среды.
На проиллюстрировано строение и разнообразие органелл, имеющих мембрану, которые обычно присутствуют в эукариотической клетке (в данном случае в типичной животной клетке). Каждая органелла содержит один или несколько компартментов. Например, эндоплазматический ретикулум (ЭПР) представляет собой один компартмент; напротив, аппарат Гольджи состоит из нескольких окруженных мембранами компартментов, обладающих определенными биохимическими функциями.
Для митохондрии характерны два компартмента, матрикс и межмембранное пространство, содержащие набор определенных макромолекул.
Цитозоль может считаться одним компартментом, ограниченным плазматической мембраной и находящимся в контакте с наружной частью мембраны всех внутриклеточных органелл. Цитоплазма состоит из цитозоля и органелл. Аналогичным образом, нуклеоплазма ограничена внутренней ядерной мембраной.
Каждая органелла содержит уникальный набор белков (как мембранных, так и растворимых), липидов и других молекул, необходимых для выполнения ее функций. Некоторые липиды и белки ковалентно связаны с олигосахаридами. По мере роста и деления клеток должны синтезироваться их новые компоненты, необходимые для роста, деления и окончательного распределения внутриклеточного материала между двумя дочерними клетками. При дифференцировке и развитии клетки, а также в ответ на воздействие таких внешних факторов, как стресс, происходит синтез компонентов органелл.
Однако компоненты не всегда образуются в той органелле, где они функционируют. Обычно различные макромолекулы образуются в сайтах, специально предназначенных для их синтеза. Например, большинство белков образуются на рибосомах цитозоля, который является оптимальной средой для функционирования рибосом и синтеза белка.
Возникает следующий вопрос: каким образом компоненты органелл попадают в места их функционирования? С начала 1970-х гг. этот вопрос являлся центральным в клеточной биологии. Как следует из рисунке ниже, существует, по крайней мере, восемь основных типов органелл, каждая из которых состоит из сотен или тысяч различных белков и липидов.
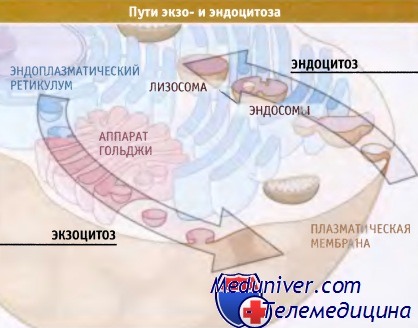 Экзоцитоз и эндоцитоз.
Экзоцитоз и эндоцитоз.
В экзоцитозе участвуют эндоплазматический ретикулум (включая ядерную оболочку)
и аппарат Гольджи (представлена одна стопка цистерн).
Эндоцитоз происходит с участием ранних и поздних эндосом и лизосом.
Все эти молекулы должны транспортироваться в органеллы, в которых они выполняют свои функции. Большинство белков образуется в цитозоле, и поэтому возникает вопрос: как они доставляются в соответствующие органеллы или же выходят из клетки, если принадлежат к секретируемым белкам? Во многих случаях ответом на этот вопрос служит наличие в молекуле белка специальных сигналов, обычно называемых сигналами сортировки или сигналами адресования. Они представляют собой короткие последовательности аминокислот, присутствующие в первичной структуре тех белков, которые должны быть локализованы не в цитозоле. Каждый адрес назначения белковой молекулы связан с одним или несколькими различными видами сигналов.
Сигналы сортировки узнаются специальными системами клетки по мере продвижения белка к месту назначения. Как показано на рисунке ниже, существует два основных транспортных механизма: экзоцитоз (или секреторный путь) и эндоцитоз, при котором материал (карго) транспортируется из клетки и в клетку соответственно.
Для всех вновь синтезированных белков, предназначенных для секреции из клетки, или для поступления в органеллы путем экзо- или эндоцитоза на мембране ЭПР существует общая точка входа. Сигналами к транслокации белка через мембрану ЭПР служат сигнальные последовательности. В настоящей главе мы рассмотрим сигналы сортировки, которые направляют белки к местам их назначения.
Находясь в ЭПР, белок не может транспортироваться через цитоплазму, и единственным путем его попадания в другие органеллы, окруженные мембранами, является везикулярный транспорт. Транспортные везикулы в основном состоят из белков и липидов и, как говорят, «отпочковываются» от мембраны. После того как везикула отпочковалась, она сливается со следующим компартментом, находящимся на ее пути. Компартмент, из которого возникла везикула, обычно называется донорский компартмент (или исходный компартмент), а компартмент назначения (или целевой) обычно называется акцепторный компартмент.
Транспортные везикулы прямым или непрямым путем переносят белки из ЭПР ко всем остальным компартментам на пути экзо- или эндоцитоза. При эндоцитозе везикулы образуются на плазматической мембране. Эти везикулы транспортируют заключенный в них материал в эндосомы, из которых образуются другие везикулы, переносящие материал в другие компартменты. Таким образом, состав транспортных везикул различается в зависимости от их происхождения и компартмента назначения.
Везикулярный транспорт создает проблему для органелл, с которыми обмениваются везикулы. Для нормального функционирования должен поддерживаться определенный внутренний состав органелл. Однако каким образом этого можно достичь, если везикулы все время изменяют этот состав? Масштаб проблемы становится очевидным при расчете эффективности транспорта. По пути эндоцитоза количество мембранных белков и липидов, эквивалентное их общему содержанию в плазматической мембране, способно транспортироваться через органеллы менее чем за час. Если сравнить со временем, необходимым для синтеза новой органеллы (обычно один день), то такая скорость впечатляет.
Решение этой проблемы связано с селективностью транспортного процесса. При отпочковывании в везикулу проходят только те белки, которые необходимо транспортировать. Резидентные белки органеллы в везикулу не попадают. Везикула удерживает эти белки и передает их следующей, находящейся на пути везикуле. Для поддержания гомеостаза между органеллами, по своей природе везикулярный транспорт всегда должен быть двунаправленным, т. е. компоненты донорского компартмента не должны непрерывно переноситься в акцепторный компартмент.
Механизм рециклирования возвращает некоторые компоненты везикул в донорский компартмент для их повторного использования. Поскольку селективность отбора белков несовершенна, специальный механизм утилизации возвращает назад резидентные белки, случайно попавшие в везикулу из донорского компартмента.
![]() При везикулярном транспорте везикула отпочковывается
При везикулярном транспорте везикула отпочковывается
от мембраны одного компартмента и сливается с мембраной другого компартмента. ![]() Пути транспорта белков в типичной животной клетке.
Пути транспорта белков в типичной животной клетке.
Почти все транспортные потоки могут осуществляться в двух направлениях.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Научная электронная библиотека

§ 3.1.4. Строение клетки
Размеры клетки широко варьируют от 0,1 мкм (некоторые бактерии) до 155 мм (яйцо страуса). У всех клеток, независимо от их формы, размеров, функциональной нагрузки обнаруживается сходное строение (рис. 3.13).
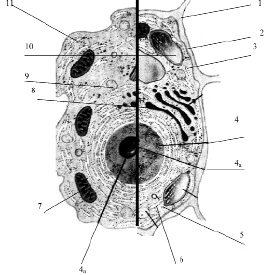
Рис. 3.13. Схема строения живой клетки: 1 – оболочка; 2 – мембрана; 3 – цитоплазма; 4 – ядро; 4а – ядрышко; 5 – рибосомы; 6 – эндоплазматическая сеть (ЭПС); 7 – митохондрии; 8 – комплекс гольджи; 9 – лизосомы; 10 – пластиды; 11 – клеточные включения
Снаружи клетка одета мембраной. Внутренняя часть клетки содержит многочисленные органоиды – структурные образования клетки, выполняющие определенные функции жизнедеятельности клетки.
1. Оболочка. Присутствует только у растительных клеток. Состоит из волокон целлюлозы. Функции оболочки: защита клетки от внешних повреждений, придает стабильную форму клетки, эластичность растительным тканям.
Повреждение наружной оболочки приводит к гибели клетки (цитолиз).
2. Мембрана. Тончайшая структура (75 Ǻ), состоит из двойного слоя молекул липидов и одного слоя белков. Такая структура обеспечивает уникальную эластичность и прочность мембране
 участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
Явление фагоцитоза – поглощение клеткой твердых частиц – впервые было описано русским врачом Мечниковым. Фагоцитарная особенность лежит в основе процесса иммунитета. Особенно развита у лейкоцитов, клеток костного мозга, лимфатических узлов, селезенки, надпочечников и гипофиза.
Пиноцитоз – поглощение клеткой растворов – состоит в том, что мельчайшие пузырьки жидкости втягиваются через образующуюся воронку, проникают через мембрану и усваиваются клеткой.
3. Цитоплазма – внутренняя среда клетки. Представляет собой гелеобразную жидкость (коллоидная система), состоит на 80 % из воды, в которой растворены белки, липиды, углеводы, неорганические вещества. Цитоплазма живой клетки находится в постоянном движении (циклоз).
 транспортировка питательных веществ и утилизация продуктов обмена клетки;
транспортировка питательных веществ и утилизация продуктов обмена клетки;
 буферность цитоплазмы (постоянство физико-химических свойств) обеспечивает гомеостаз клетки, поддерживает постоянные нужные параметры жизнедеятельности;
буферность цитоплазмы (постоянство физико-химических свойств) обеспечивает гомеостаз клетки, поддерживает постоянные нужные параметры жизнедеятельности;
 поддержание тургора (упругость) клетки;
поддержание тургора (упругость) клетки;
 все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
4. Ядро – обязательный органоид эукариотических клеток. Впервые было исследовано и описано Р. Броуном в 1831 г. В молодых клетках расположено в центре клетки, в старых – смещается в сторону. Снаружи ядро окружено мембраной с крупными порами, способными пропускать крупные макромолекулы. Внутри ядро заполнено клеточным соком – кариоплазмой, основная часть ядра заполнена хроматином – ядерным веществом, содержащим ДНК и белок. Перед делением хроматин образует палочковидные хромосомы. Причём, хромосомы одинакового строения (но содержащие разные ДНК!) образуют пары, зрительно воспринимаемые как одно целое (рис. 3.14).

Рис. 3.14. Хромосомный набор человеческой клетки перед началом деления
Структурирование всех хромосом в пары свидетельствует о том, что число хромосом – чётное. Поэтому, его часто обозначают 2n, где n – количество хромосомных пар, а соответствующий набор хромосом называют диплоидным. Например, у голубей n = 40 (80 хромосом), у мухи n = 6 (12 хромосом), у собаки n = 39 (78 хромосом), у аскариды n = 1 (2 хромосомы). У человека n = 23 (46 хромосом). Однако, в половых клетках число хромосом в два раза меньше. Поэтому набор хромосом в половых клетках называется гаплоидным. Клетки, не являющиеся половыми называются соматическими. Иногда клетки с гаплоидным набором хромосом называют гаплоидными клетками, а с диплоидным набором хромосом – диплоидными клетками.
При слиянии двух родительских гаплоидных половых клеток образуется диплоидная клетка, дающая начало новому организму с набором генов отца и матери
Совокупность всех хромосом ядра (а значит и генов) клетки называется генотип. Именно генотип определяет все внешние и внутренние признаки конкретного организма.
В соматических клетках 44 Х-образные хромосомы (22 пары) у женщин и мужчин идентичны (сходны по строению), их называют аутосомами. А 23-я пара имеет конфигурацию ХХ – у женщин и ХY – у мужчин. Эти пары хромосом именуются половыми хромосомами.
В половых клетках 22 хромосомы также одинаковые у яйцеклеток и у сперматозоидов, а 23-я хромосома конфигурации Х – у яйцеклетки и Х или Y – у сперматозоидов. Поэтому при слиянии половых клеток и образовании пар хромосом, 23-я пара будет ( <ХY>или <ХХ>) определять пол будущего ребенка.
Необходимо помнить, что хотя в соматических клетках набор хромосом диплоидный (2n), однако, перед началом деления клеток происходит репликация ДНК, то есть, удвоение их количества, а, значит, и удвоение
количества хромосом. Поэтому перед началом деления соматической клетки в ней насчитывается 4n хромосом (рис. 16). Она становится тетраплоидной.
– хранение генетической информации;
– контроль за всеми процессами, происходящими в клетке: делением, дыханием, питанием и др.
4а. Ядрышко – структура, содержащаяся в ядре. Ядро может содержат 1, 2 или более ядрышек. Функция ядрышка – формирование рибосом.
Следует отметить, что не все клетки имеют оформленное ядро. Клетки, имеющие ядро называются эукариотическими или эукариотами. Клетки, не имеющие ядра, называются прокариотическими или прокариотами. Функции ядра у прокариот несёт одна нить ДНК (именуется хромосома), в которой хранится вся генетическая информация. К прокариотам относятся бактерии и сине-зеленые водоросли. Как правило, у прокариотов отсутствуют и некоторые другие органоиды. Размеры прокариотических клеток меньше, чем размеры эукариот.
5. Рибосомы – самые мелкие органоиды клетки. Были обнаружены в 1954 г. Французским ученым Паладом. Рибосомы были обнаружены в цитоплазме, а также на гранулярной ЭПС и в ядре.
Функция рибосом: обеспечение биосинтеза белка.
6. Эндоплазматическая сеть. Представляет собой каналы и полости, ограниченные мембраной. Различают две разновидности ЭПС: гранулярная ЭПС и агранулярная ЭПС. Гранулярная ЭПС морфологически отличается от агранулярной наличием на ее поверхности многочисленных рибосом (на агранулярной ЭПС рибосомы отсутствуют).
Функции эндоплазматической сети:
– участие в синтезе органических веществ: на гранулярной ЭПС синтезируются белки, на агранулярной – липиды и углеводы;
– транспортировка продуктов синтеза ко всем частям клетки.
Несложно уяснить, что гранулярная ЭПС характерна для клеток, синтезирующих белки (например клетки желез внутренней секреции), агранулярная ЭПС характерна для клеток-производителей углеводов и липидов (например клетки жировой ткани).
7. Митохондрии – крупные органоиды, состоящие из двойного слоя мембран: наружная – гладкая, внутренняя образует многочисленные гребнеобразные складки – кристы. Внутри митохондрии заполнены жидкостью (матрикс).
Функции митохондрий: основная функция митохондрий – обеспечение клетки энергией. Этот процесс происходит за счет синтеза аденозинтрифосфорной кислоты (АТФ) (рис. 3.15), в которой фрагмент

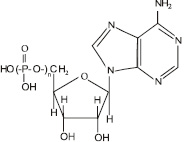
Рис. 3.15. Структурная формула аденозинфосфорных кислот. Для аденозинтрифосфорной кислоты n = 3, для аденозиндифосфорной кислоты n = 2, для аденозинмонофосфорной кислоты n = 1
При взаимодействии молекулы аденозинтрифосфорной кислоты с водой отщепляется один остаток фосфорной кислоты, в результате чего образуется аденозиндифосфорная кислота – АДФ и выделяется огромное количество энергии:
АТФ + Н2О = АДФ + Н3РО4 + 10 000 калорий.
Впоследствии от АДФ может отщепляться еще один остаток фосфорной кислоты, образуя АМФ – аденозинмонофосфорную кислоту.
АДФ + Н2О = АМФ + Н3РО4 + 10 000 калорий[37].
Освободившаяся энергия используется для жизнедеятельности клетки (КПД процесса превышает 80 %!).
Наряду с распадом АТФ и выделением энергии в клетке постоянно происходит синтез АТФ и накопление энергии (обратные реакции).
Количество митохондрий в клетке зависит от потребности последней в энергии. Так, в клетках кожи человека находится в среднем 5–6 митохондрий, в клетках мышц – до 1000, в клетках печени – до 2500!
8. Комплекс Гольджи. Итальянский ученый Гольджи обнаружил и описал структуру клетки, напоминающую стопки мембран, цистерны, пузырьки и трубочки. Расположена эта система чаще всего возле ядра.
Функции комплекса Гольджи: в полостях комплекса накапливаются всевозможные продукты обмена клетки, которые по каким-либо причинам не вывелись наружу. В последствии эти продукты могут быть использованы клеткой для процессов жизнедеятельности. Из пузырьков и цистерночек комплекса Гольджи в растительных клетках образуются вакуоли, заполненные клеточным соком.
9. Лизосомы – мелкие органоиды. Представляют собой пузырьки, окруженные мембраной. Внутри лизосомы заполнены пищеварительными ферментами (обнаружено 12 ферментов), которые расщепляют и переваривают крупные макромолекулы (белки, полисахариды, нуклеиновые кислоты).
Функции лизосом: растворение и переваривание макромолекул. Лизосомы участвуют в фагоцитозе. Понятно, что основная функция по перевариванию поступающих в клетку частиц принадлежит лизосомам.
10. Пластиды. Эти органоиды характерны только для растительных клеток. Форма напоминает двояковыпуклую линзу. Структура пластид напоминает таковую у митохондрий: двойной слой мембраны. Наружная – гладкая, внутренняя образует складки, называемые тилакоидами. На тилакоидах происходит основной жизненно важный для всех зеленых растений процесс – фотосинтез:

Пластиды бывают трех типов:
1) Хлоропласты – зеленые пластиды. Их цвет обусловлен наличием хлорофилла. Хлорофилл – основное вещество хлоропластов (имеет зеленый цвет). Только благодаря хлорофиллу возможен процесс фотосинтеза (см. раздел 4.2). Хлоропласты придают зеленый цвет растительным организмам.
2) Хромопласты – пластиды, имеющие различные окраски: от ярко-желтого до пурпурно-багряного. Наличие различных пигментов окрашивают плоды, цветки и осенние листья растений в соответствующие цвета. Этот факт особенно важен для привлечения насекомых к цветкам, как природный индикатор созревания плодов и др.
3) Лейкопласты – бесцветные пластиды, в которых происходит накопление запасных питательных веществ (например, крахмала).
Некоторые виды пластид могут переходить друг в друга: например, переход хлоропластов в хромопласты: созревание томатов, яблок, вишни, и т. д.; изменение окраски листьев в осенний период времени. Лейкопласты могут переходить в хлоропласты: позеленение картофеля на свету. Это доказывает общность происхождения пластид.
11. Клеточные включения. Вакуоли. Это непостоянные и необязательные составляющие клетки. Они могут появляться и исчезать в течение всей жизни клетки. К ним относятся капли жира, зерна крахмала и гликогена, кристаллы щавелево-кислого кальция и др. Жидкие продукты обмена называются клеточным соком и накапливаются они в вакуолях. В клеточном соке растворены сахара, минеральные соли, пигменты и т. д. Чем старше клетка, тем больше клеточного сока накапливает клетка. Молодые клетки практически не содержат вакуолей.
Помимо перечисленного некоторые специализированные клетки обладают специальными органоидами. К ним относятся:
– реснички и жгутики, представляющие собой выросты мембраны клетки, осуществляющие движения клетки. Они имеются у одноклеточных организмов и многоклеточных (кишечный эпителий, сперматозоиды, эпителий дыхательных путей);
– миофибриллы – тонкие нити мышечных клеток, участвующие в сокращении мышц;
– нейрофибриллы – органоиды, характерные для нервных клеток и участвующие в проведении нервных импульсов. Кроме того, в состав клеток входят центриоли – две (иногда более) цилиндрические структуры диаметром около 0,1 мкм и длиной 0,3 мкм. Место расположения центриолей в период между делениями клетки считается серединой клеточного центра. При делении клетки центриоли расходятся в противоположные стороны – к полюсам, определяя ориентацию веретена деления (рис. 16).
Следует иметь в виду, что, хотя животные и растительные клетки имеют много общего, но между ними существуют и серьёзные различия (табл. 3.1).
Более общая классификация клеток представлена на рис. 3.16.
Одно из основных отличий бактерий от архей, состоит в химическом составе мембраны. Бактерии отделены от внешней среды двойным слоем липидов (жиров и жироподобных веществ). Мембраны архей состоят из терпеновых спиртов.
Чем представлена транспортная система клетки
• Существует два основных типа мембранных транспортных белков: каналы и переносчики
• Ионные каналы катализируют быстрый и избирательный транспорт ионов по электрохимическому градиенту
• Транспортеры и насосы представляют собой белки-переносчики, которым необходима энергия для транспорта веществ против электрохимического градиента
• В каждой клетке несколько различных мембранных транспортных белков действуют как единое целое
Некоторые из транспортных белков находятся в плазматической мембране, в то время как другие входят в состав мембраны клеточных органелл. Для поддержания определенного состава внутриклеточной среды и клеточных компартментов необходимо, чтобы транспортные белки обладали избирательностью по отношению к определенным метаболитам. В зависимости от характера процесса транспорта мембранные транспортные белки подразделяются на две группы: каналы и переносчики.
Канальные белки содержат поры, через которые при открытом канале с высокой скоростью проходят метаболиты. Белок-переносчик связывает метаболит, находящийся по одну сторону мембраны, после чего конформация его меняется, и метаболит высвобождается с другой стороны мембраны.
Существует несколько типов белков, образующих каналы в мембранах. Порины, присутствующие в клетках некоторых прокариот, в митохондриях и в нексусах, где соединяется цитоплазма соседних клеток, обеспечивают прохождение метаболитов в соответствии с размером их молекул.
К более селективным каналам относятся комплексы ядерных пор и каналы, обеспечивающие перемещение белков через мембрану эндоплазматического ретикулума. В статьях на сайте будут рассмотрены ионные каналы и аквапорины, через которые осуществляется селективный трансмембранный перенос ионов и молекул воды соответственно. К настоящему времени описано более 100 различных типов канальных белков. Все они обладают следующими характеристиками:
• селективностью по отношению к переносимому метаболиту,
• высокой скоростью переноса метаболита и
• воротным механизмом, регулирующим транспорт метаболитов.
Участок канального белка, через который осуществляется трансмембранный транспорт, называется порой канала. Канал может находиться в нескольких различных конфигурациях. Некоторые каналы состоят из одного белка, трансмембранные сегменты которого образуют пору. Другие существуют в виде олигомеров, состоящих из идентичных или разных субъединиц, которые вместе образуют пору.
Наконец существуют каналы, состоящие из двух и более субъединиц, каждая из которых сама образует пору. Часто комплексы олигомеров каналов регулируются или позиционируются на определенных мембранах с участием дополнительных субъединиц.
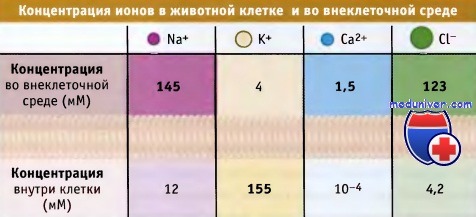
Большинство канальных белков обладают высокой селективностью в отношении определенных метаболитов, например ионов натрия (Na+), калия (К+), кальция (Са2+), хлорида (Сl-) или воды. Другие канальные белки являются неселективными катионными или анионными каналами. Как будет показано далее, при рассмотрении свойств специфических канальных белков, их поры характеризуются определенными структурными особенностями, т. е. являются селективным фильтром, который позволяет им проявлять избирательность по отношению к различным метаболитам.
![]() Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Растворенные метаболиты с максимальной скоростью диффундируют через пору каналов,
а переносчики связывают метаболит с одной стороны мембраны, затем претерпевают конформационные изменения и высвобождают его с другой стороны.
Этот процесс протекает значительно медленнее.
Направление потока ионов растворенных веществ через канал определяется электрохимическим градиентом. Иными словами, метаболиты движутся через канал в энергетически-выгодном направлении, т. е. по электрохимическому градиенту. Например, для покоящейся клетки, схематически изображенной на рисунке ниже, существует направленный в клетку ионный поток через Na+-, Са2+-, Cl—каналы, и противоположный, проходящий через К+-каналы.
Так как других источников энергии, кроме электрохимического градиента, в данном случае не используется, то такой тип транспорта называется пассивным. Канальные белки обеспечивают высокую скорость транспорта. Так, для ионных каналов скорость составляет 108 в 1 с, что близко к максимальной скорости диффузии ионов в водной среде.
Транспортные мембранные белки контролируют различные клеточные функции при участии воротного механизма. В основе этого механизма лежит индукция конформационных изменений в белке в ответ на воздействие определенного стимула. Эти изменения приводят к открытию или закрытию канала. Например, открытие и закрытие ионных каналов могут регулироваться лигандами, величиной электрического потенциала, механическими или температурными воздействиями. Активация каналов может происходить очень быстро, что делает их идеальным инструментом обработки поступающих в клетку сигналов.
Например, передача нервного импульса зависит от электрического заряда, который создается ионным каналом в мембране. Каналы также играют важную роль в регуляции объема клетки и внутриклеточного pH, в транспорте солей и воды через эпителиальные клетки, в закислении внутриклеточных органелл и во внеклеточной передаче сигналов.
 Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
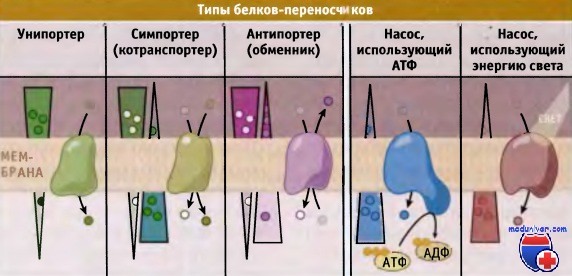
Транспортеры делятся на три типа, каждый из которых осуществляет один из типов переноса: унипорт, симпорт, или антипорт.
Показано направление электрохимического градиента через мембрану.
В зависимости от типа белка-переносчика транспорт может происходить или в направлении градиента (от высокой концентрации к низкой), или против него.
При транспорте субстратов против градиента концентрации, белки-переносчики используют свободную энергию электрохимического градиента, АТФ или других источников. Поскольку при этом происходит потребление энергии, процесс носит название активный транспорт. Белки-переносчики подразделяются на две группы, транспортеры и насосы.
Для транспортировки субстратов через мембрану транспортеры используют энергию электрохимического градиента. Они подразделяются на унипортеры, симпортеры (или котранс-портеры) и антипортеры (обменники). Для энергозависимого транспорта метаболитов мембранные насосы используют энергию, например высвобождающуюся при гидролизе АТФ. По сравнению с канальными белками, переносчики характеризуются более медленной скоростью транспорта, порядка 1000 молекул в 1 с.
Различают два типа активного транспорта, первичный и вторичный. Белки-переносчики, осуществляющие первичный активный транспорт, в качестве источника энергии используют АТФ и транспортируют метаболиты против электрохимического градиента. Таким образом, они помогают поддерживать градиент концентрации метаболитов по обеим сторонам мембран клетки. Са2+-АТФаза и Na+/К+-АТФаза являются характерными примерами транспортных насосов, которые управляют процессом первичного активного транспорта.
Белки-переносчики, которые участвуют во вторичном активном транспорте, непосредственно не используют энергию АТФ. Вместо этого они используют свободную энергию электрохимического градиента, который возникает за счет первичного активного транспорта. Вторичный активный транспорт происходит с участием симпортеров и антипортеров. Механизмы транспорта метаболитов представлены на рисунке ниже.
В клетке все типы мембранных транспортных белков-каналы, транспортеры и насосы-функционируют совместно. Таким образом, работа какого-либо одного типа транспортного белка зависит от работы других. В данной главе мы рассмотрим несколько примеров такого взаимодействия транспортных белков. Например, ионный градиент по обеим сторонам мембраны поддерживается за счет сложных взаимодействий между различными типами транспортных белков. Для правильного функционирования эпителиальных клеток, например почек, слизистой кишечника и легкого, необходимо протекание транспортных процессов с участием различных ионов и метаболитов. Мы также рассмотрим роль нарушений процессов транспорта в развитии различных заболеваний.
![]() Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021



